
El efecto alelopático que tienen los extractos de Schinus molle, Eucalyptus camaldulencis y Festuca rubra, sobre la especie Phaseolus vulgaris.
RESUMEN
La alelopatía juega un importante papel en el mecanismo de dominación vegetal. Hay varios métodos para evaluar el potencial alelopático de la plantas. Se quiso comprobar los efectos alelopáticos que tienen las especies de Eucalyptus camaldulencis, Schinus molle y Festuca rubra en las muestras con pasto, seguidas por las muestras con E. camaldulensis y finalmente se obtuvieron resultados contrarios a los esperados en las muestras con S. molle. Llegando a la conclusión de que la Festuca rubra contiene sustancias alelopáticas más potentes que el E. camaldulencis, y por el contrario, el S. molle parece no tener este tipo de propiedades.
INTRODUCCIÓN
Las interacciones planta- planta son diversas y se establecen entre aquelloss vegetales que comparten el mismo hábitat; aquellas que son mediadas por la presencia de metabólicos secundarios, se refieren al fenómeno llamado alelopatía. El término fue propuesto por Molisch en 1973 y se deriva de dos raíces griegas, allelon= recíproco pathos = sufrimiento (Vázquez y Orozco, 1993). Se definió a la alelopatía como cualquier proceso que involucra la producción de metabólicos secundarios producidos por plantas y microorganismos, los cuales influyen sobre otors sistemas biológicos.
La competencia por interferencia los individuos afectan directamente a otros por medios físicos, o químicos.La competencia química (alelopatía) ocurre cuando un organismo produce una toxina que actúa a cierta distancia después de difundirse a través del ambiente (Vázquez y Orozco, 1993).
Algunos organismos, principalmente las plantas, suelen estar limitados en cuanto a su distribución por “venenos”, “antibióticos” o agentes alelopaticos. (Krebs, 1985). la producción de toxinas para inhibir el crecimiento de las plantas vecinas parece una estrategia evolutiva conveniente para las plantas.
Los efectos alelopáticos de Festuca rubra sobre los cultivos no está tan estudiado y actualmente está en discusión. Sin embargo, hay algunos casos particulares que involucran el efecto de este sobre comunidades de plantas. Gran cantidad de pastos alelopáticos son bien conocidos, estos en su gran mayoría producen y liberan triterpenos que son especialmente activos contra las dicotiledóneas (Caldiz y Fernández 1999).
En cuanto al E. camaldulencis, se ha demostrado que los residuos frescos de las hojas, son efectivos en la reducción del crecimiento de malezas. Contiene terpenoides que se liberan en la atmósfera y afectan a organismos vecinos.
El eucalipto es un árbol de la familia de las mirtáceas, de tronco derecho y copa por lo regular cónica, que puede crecer hasta 100 metros de altura (Abellan, 1980). Eucalyptus camaldulencis. Dehnh o Eucalipto rojo. Es un árbol alto que alcanza hasta los 40 m (raras veces), con tronco corto y copa extendida. Corteza lisa, que se desprende en forma de escamas o tiras, moteada de ante y gris. Una de las especies más variables y extendidas de Australia;(Humphries, 1982) es uno de los árboles que más extensamente se cultivan en las plantaciones de eucaliptos del mundo. Se suele plantar en suelos pesados y en Españña es particularmente estimado por su madera para hacer puntuales en minas y como combustible. Fue introducido en la región mediterránea a principios del siglo XIX.
El Schinus molle. L.Molle, pimentero falso, Turbinto
Es un arbusto o árbol perennifolio de hasta 12 m de altura (Humphries, 1982). Ramas esbeltas colgantes, muy aromático al ser estrujado. Nativo de las montañas de América del Sur, plantado como ornamental en el sur de Europa y más o menos naturalizado en algunas localidades.
El S. molle es una especie intruducida en México desde Perú durante el siglo XVI. Se adaptó tan perfectamente a las condiciones del altiplano mexicano que ahora es prácticamente el único árbol que se puede encontrar sobre enormes extensiones dedicadas a la agricultura en valles como el de Puebla y Pachuca. Es posible que esta planta haya sido en parte responsable de la desaparición de la flora arbórea nativa de estas regiones, ya que sus semillas son eficientemente dispersadas por algunas aves migratorias que regurgitan las semillas después de que han solubilizado los azúcares que las cubren. Al germinar y establecerse las semillas, el pirú acaba sustituyendo al árbol en cuya base germinó.(Vázquez, y Orozco, 1996).
El Phaseolus vulgaris es cultivado en una gran parte de América Latina convirtiéndose en uno de los cultivos de mayor importancia.
La germinación de las semillas comprende tres etapas sucesivas que se superponen parcialmente 1) la absorción del agua por imbibición, causando su hinchamiento y la ruptura final de la testa; 2) El inicio de la actividad enzimática y del metabolismo respiratorio, translocación y asimilación de las recervas alimentarias en las regiones en crecimiento del embrión; 3) El crecimiento y la división celular que provoca la emergencia de la radícula y posteriormente de la plúmula. En la mayoría de las semillas el agua penetra inicialmente por el micrópilo y la primera manifestación de la germinación exitosa es la emergencia de la radícula. (Nava y Anaya, 2003).
OBJETIVOHIPÓTESIS
Comprobar el efecto alelopático que tiene los extractos de Schinus molle, Eucalyptus camaldulencis y Festuca rubra, sobre la especie Phaseolus vulgaris. Para verificar las propoedades alelopáticas que se le atribuyen a estas plantas.
Si tenemos en cuenta que las especies de Schinus molle, Eucalyptus camaldulencis y Festuca rubra, se les atribuyen propiedades alelopáticas, deben inhibir el crecimiento de Phaseolus vulgaris.
DISEÑO EXPERIMENTAL:
La variable independiente son los extractos de que usaremos de Schinus molle, Eucalyptus camaldulencis y Festuca rubra en Phaseolus vulgaris.
La variable dependiente es el crecimiento de Phaseolus vulgaris.
Tuvimos que volver a repetir el experimento, debido a que no cuidamos la concentración que debía tener cada extracto, lo que podría provocar considerables irregularidades en los resultados. Además de que las semillas no germinaron y se secaron, por lo que no se pudo obtener ningún resultado útil para el experimento.
Cuando repetimos el experimento, los extractos se hicieron con 250 mL de agua y 50g de hojas trituradas de la planta correspondiente, esto se llevó a cabo con la licuadora; después se filtraron con un paño limpio.
MATERIAL Y MÉTODO
Se utilizaron tres platos de unicel con algodón, cada uno con la misma cantidad, como testigos y otros tres como experimentales para cada una de las sustancias, cada uno de estos platos tenía nueve especímenes de P. vulgaris acomodados en tres hileras de tres.
A cada lote experimental se le agregó 2 mL de sustancia alelopática correspondiente por cada semilla, mientras que a los testigos sólo se les agregó agua, la suficiente como para que el algodón se mantuviera húmedo. Los líquidos se manejaron con pipetas y propipetas, para tener una concentración exacta. Una vez concluido, se envolvían con plástico para brindarle protección a las semillas, éste se perforaba medio centímetro, para permitir la libre circulación de aire.
RESULTADOS
En el caso del E. camaldulensis se obtuvieron los siguientes resultados:

Eu(n) son los lotes experimentales.
EuC(n) son los lotes testigos.
Aquí se observa la cantidad de especímenes de P. vulgaris que germinaron por cada lote, tanto experimental como testigo, y el tamaño que alcanzaron. En el caso de los lotes testigos, el máximo tamaño alcanzado fue de 2.5 cm; y en el caso de los lotes experimentales el máximo tamaño alcanzado fue de 2.1 cm. EuC(n) son los lotes testigos.
En el caso de F. rubra se obtuvieron los siguientes resultados

P(n) son los lotes experimentales.
PC(n) son los lotes testigos.
Aquí se observa la cantidad de especímenes de P. vulgaris que germinaron por cada lote, tanto experimental como testigo, y el tamaño que alcanzaron. En el caso de los lotes testigos, el máximo tamaño alcanzado fue de 7.8 cm; y en el caso de los lotes experimentales el máximo tamaño alcanzado fue de 3.9 cm. PC(n) son los lotes testigos.
En el caso de S. molle se obtuvieron los siguientes resultados.
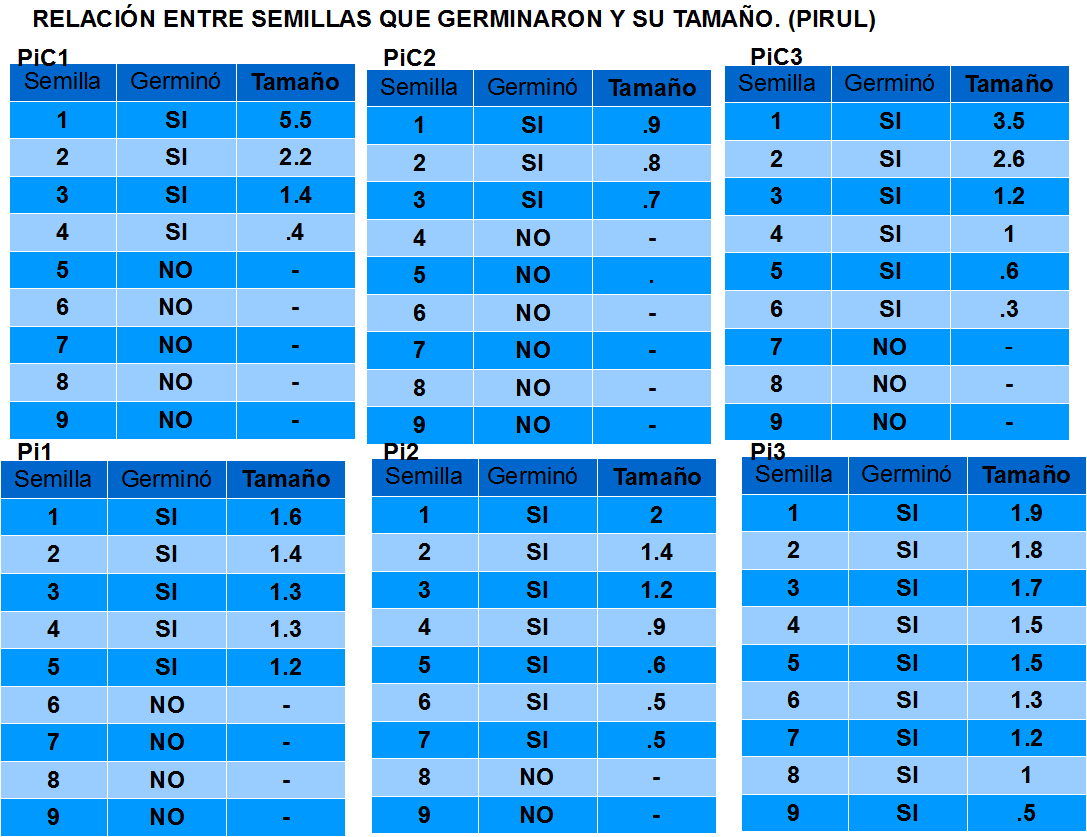
Pi(n) son los lotes experimentales.
PiC(n) son los lotes testigos.
Aquí se observa la cantidad de especímenes de P. vulgaris que germinaron por cada lote, tanto experimental como testigo, y el tamaño que alcanzaron. En el caso de los lotes testigos, el máximo tamaño alcanzado fue de 5.5 cm; y en el caso de los lotes experimentales el máximo tamaño alcanzado fue de 2 cm. PiC(n) son los lotes testigos.
Se sacó un promedio del tamaño de todos los lotes testigos y de todos los lotes experimentales, para poder observar el crecimiento relativo de cada uno.

No todas las semillas germinaron, por lo que se sacó un porcentaje para determinar si la germinación estaba relacionada con la ausencia o presencia de agentes alelopáticos.

Se midieron los retoños y se tomó en cuenta el de mayor tamaño de cada tipo de lote para comparar los que tenían agente alelopático y los que no.

DISCUSIÓN Y CONCLUSIONES
Aparentemente en cuanto a la relación de tamaño máximo alcanzado por los lotes, los que tenían agentes alelopáticos presentes alcanzaron un mayor tamaño que los que no tenían. Sin embargo en cuanto al promedio de crecimiento, el S. molle tiene un efecto inverso al esperado, y en los lotes con agente alelopático germinaron más semillas de P. vulgaris que los que carecían de este. Esto no necesariamente se puede deber a la ineficiencia de el S. molle como agente alelopático, de lo contrario todos los gráficos habrían salido iguales, pero en el caso de tamaño máximo alcanzado por cada lote, el experimental creció menos que el testigo. Esta diferencia radica en cuanto a la germinación, no en cuanto al crecimiento de las semillas. Por lo que no se puede descartar completamente al S. molle como agente alelopático. En cuanto al F. rubra , fue el que arrojó resultados más notables, en cuanto a tamaño máximo alcanzado, fue el que obtuvo el mayor resultado. También en cuanto al indice de germinación. Lo mismo sucede en cuanto a los promedios de crecimiento. Lo que nos da como conclusión que el F. rubra contiene agentes alelopáticos muy efectivos. En segundo lugar se encuentra el E. camaldulensis, cuyos resultados fueron los esperados, un mayor índice de germinación en lotes testigos que en experimentales, al igual que un mayor tamaño y un promedio de crecimiento mayor.
Aparentemente en cuanto a la relación de tamaño máximo alcanzado por los lotes, los que tenían agentes alelopáticos presentes alcanzaron un mayor tamaño que los que no tenían. Sin embargo en cuanto al promedio de crecimiento, el S. molle tiene un efecto inverso al esperado, y en los lotes con agente alelopático germinaron más semillas de P. vulgaris que los que carecían de este. Esto no necesariamente se puede deber a la ineficiencia de el S. molle como agente alelopático, de lo contrario todos los gráficos habrían salido iguales, pero en el caso de tamaño máximo alcanzado por cada lote, el experimental creció menos que el testigo. Esta diferencia radica en cuanto a la germinación, no en cuanto al crecimiento de las semillas. Por lo que no se puede descartar completamente al S. molle como agente alelopático. En cuanto al F. rubra , fue el que arrojó resultados más notables, en cuanto a tamaño máximo alcanzado, fue el que obtuvo el mayor resultado. También en cuanto al indice de germinación. Lo mismo sucede en cuanto a los promedios de crecimiento. Lo que nos da como conclusión que el F. rubra contiene agentes alelopáticos muy efectivos. En segundo lugar se encuentra el E. camaldulensis, cuyos resultados fueron los esperados, un mayor índice de germinación en lotes testigos que en experimentales, al igual que un mayor tamaño y un promedio de crecimiento mayor.
BIBLIOGRAFÍA
-Abellan, E. (1980). El eucalipto. Cultivo y aprovechamiento. Madrid: Sintes. 5.
-Caldiz, D. y Fernández, L. (1999). Recent advances in allelopathy: A science for the future. Cádiz: Universidad de Cádiz. 3, 454 y 457.
-Humphries, C. (1982). Guía de los árboles de España y de Europa. España: Omega. 226 y 276.
-Krebs, C. (1985). Ecología. Estudio de la distribución y la abundancia. México: Harla. 59 y 63.
-Nava, V. y Anaya, A. (2003).Leguminosas: Caracteristicas quimicas y su importancia en las interacciones con otros organismos. México: UAM. 35-37.
-Ricklefs, R. (2001). Invitacion a la ecología: la economía de la naturaleza. España: Editorial médica Panamericana. 455, 456 y 692.
-Vázquez, C. y Orozco, A. (1993). La destrucción de la naturaleza. México: Fondo de Cultura Económica. Capítulo III.
No hay comentarios:
Publicar un comentario